CRISPR systems are adaptable immune mechanisms which are present in many bacteria to protect themselves from foreign nucleic acids, such as viruses, transposable elements or plasmids. Recently, the CRISPR/Cas9 (CRISPR-associated protein 9 nuclease, UniProtKB/Swiss-Prot entry Q99ZW2) system from S. pyogenes has been adapted for inducing sequence-specific double stranded breaks and targeted genome editing. This system is unique and flexible due to its dependence on RNA as the moiety that targets the nuclease to a desired DNA sequence and can be used induce indel mutations, specific sequence replacements or insertions and large deletions or genomic rearrangements at any desired location in the genome. In addition, Cas9 can also be used to mediate upregulation of specific endogenous genes or to alter histone modifications or DNA methylation.
CRISPR/Cas9 polyclonal antibody

Polyclonal antibody raised in rabbit against the Cas9 nuclease (CRISPR-associated protein 9) using a recombinant protein.
Western blot was performed using HCT116 DKO cells transduced with Krab-dCas9 (2) or non-transduced (1) cells. Then, 100,000 cells were lysed in sample buffer 2x and boiled 5 min at 95°C before loading in a 15% acrylamide gel. The same sample was loaded 3x in the same gel. The membrane was cut in 3 parts for each antibody. Membrane was blocked 1h with 3% BSA at RT. Antibodies were diluted 1:1,000 in 3% BSA and incubated overnight at 4°C. Secondary incubation was done for 1h at RT (1:10,000 dilution). Anti-hnRNPA1 was used as a loading control.

| Lot | A2508-004 |
|---|---|
| Concentration | Not determined |
| Species reactivity | Streptococcus pyogenes |
| Type | Polyclonal, ChIP grade, ChIP-seq grade |
| Purity | Whole antiserum from rabbit containing 0.05% azide. |
| Host | Rabbit |
| Storage Conditions | Store at -20°C; for long storage, store at -80°C. Avoid multiple freeze-thaw cycles. |
| Precautions | This product is for research use only. Not for use in diagnostic or therapeutic procedures. |
| Applications | Suggested dilution | References |
|---|---|---|
| ChIP/ChIP-seq * | 2-5 µl/ChIP | Fig 1, 2 |
| Western Blotting | 1:5,000 | Fig 3 |
| Immunoprecipitation | 1 µl/IP | Fig 4 |
| Immunofluorescence | 1:1,000 | Fig 5 |
* Please note that the optimal antibody amount per IP should be determined by the end-user. We recommend testing 1-10 µl per IP.
TESTIMONIALI used ChIP-qPCR with the Diagenode CRISPR/Cas9 polyclonal antibody to successfully show that Cas9 binds to the target region of my sgRNA, validating my CRISPR experiment. The antibody produced minimal background signal at non-specific genomic regions. I am now using the antibody to validate further sgRNA in different CRISPR cell lines.
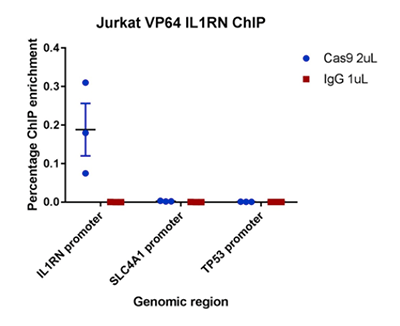
ChIP was performed on Jurkat cells expressing dCas9-VP64-mCherry and a sgRNA targeting the IL1RN promoter. Each IP was performed using 4 million cells and 2 µL CRISPR/Cas9 polyclonal antibody (Diagenode C15310258) or 1 µg rabbit IgG control antibody (Diagenode C15410206). qPCR was carried out on undiluted ChIP DNA using SYBR green and PCR primers directed against the sgRNA binding site at IL1RN, as well as two non-target regions at the SLC4A1 and TP53 promoters. ChIP enrichment was measured using the percent input method.
Researcher from the University of Manchester
- Validation data
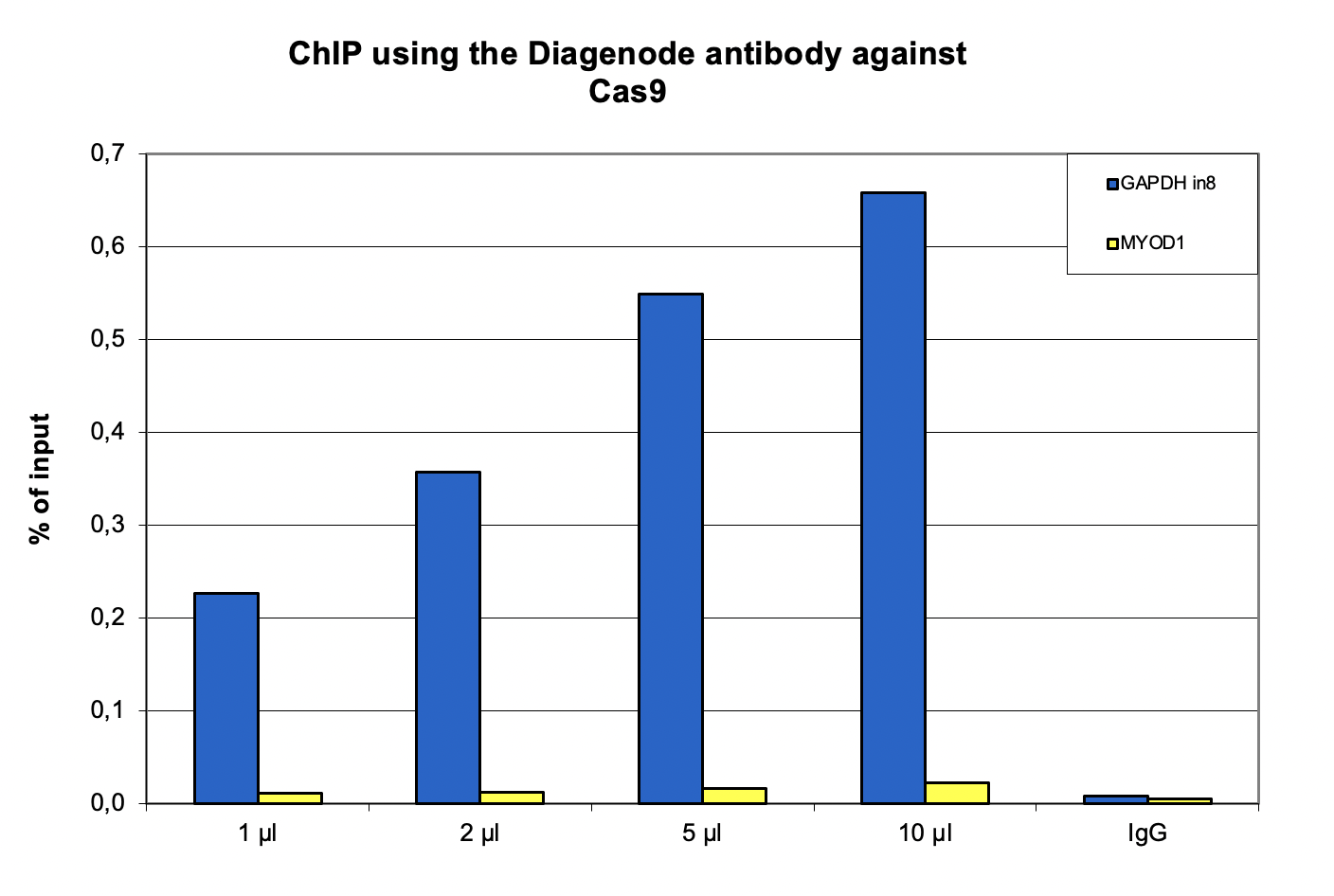
Figure 1. ChIP using the Diagenode antibody directed against Cas9
ChIP was performed on sheared chromatin from 4 million HEK293T cells stably expressing nuclease dead Cas9 and sgRNA targeting a sequence in intron 8 of the GAPDH gene, using the iDeal ChIP-seq kit for transcription factors. A titration consisting of 1, 2, 5 and 10 µl of the Diagenode antibody against Cas9 (cat. No. C15310258) was tested. IgG (2 µg/IP) was used as negative IP control. qPCR was performed with primers specific for the targeted sequence in the GAPDH gene, and for the MYOD1 gene, used as negative control. Figure 1 shows the recovery, expressed as a % of input (the relative amount of immunoprecipitated DNA compared to input DNA after qPCR analysis).A. 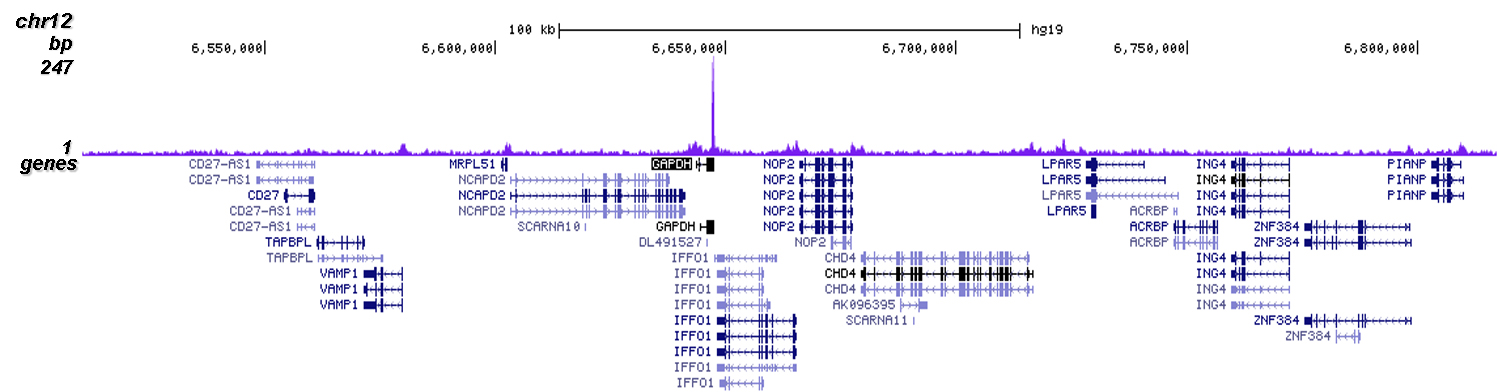
B. 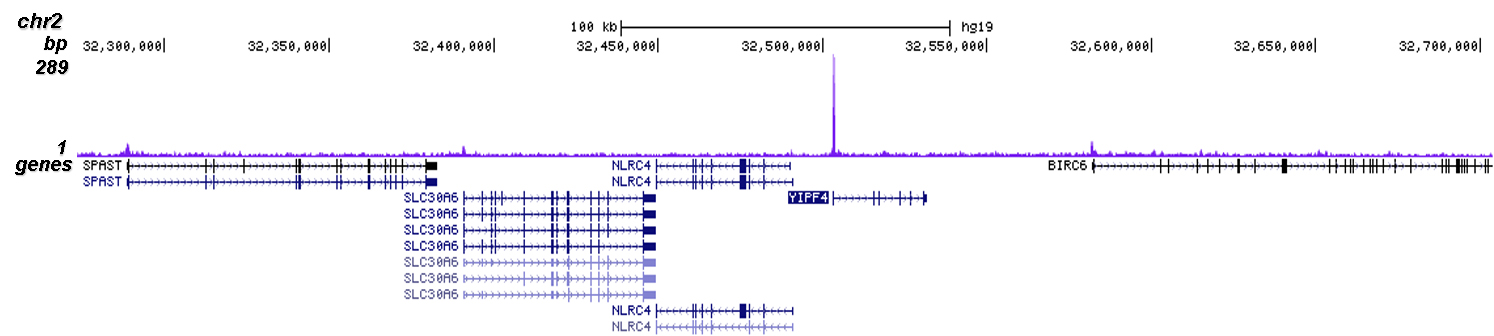
Figure 2. ChIP-seq results obtained with the Diagenode antibody directed against Cas9
ChIP was performed on sheared chromatin from 4 million HEK293T cells stably expressing dCas9 and a GAPDH sgRNA cells using 2 µl of the Diagenode antibody against Cas9 (cat. No. C15310258) as described above. The IP'd DNA was subsequently analysed on an Illumina NovaSeq. Library preparation, cluster generation and sequencing were performed according to the manufacturer's instructions. The 50 bp tags were aligned to the human genome using the BWA algorithm. Figure 2 shows the ChIP-seq profile in a region of chromosome 12 surrounding the GAPDH gene (fig 2B) and in a region of chromosome 2 surrounding an off-target peak in the YIPF4 gene.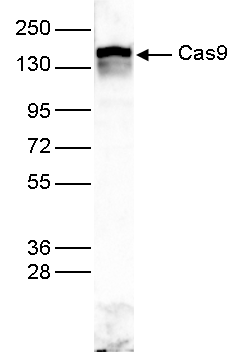
Figure 3. Western blot analysis using the Diagenode antibody directed against Cas9
Western blot was performed on protein extracts from HEK293 cells transfected with dCas9 using the Diagenode antibody against CRISPR/Cas9 (cat. No. C15310258). The antibody was diluted 1:5,000. The marker is shown on the left, the position of the Cas9 protein is indicated on the right.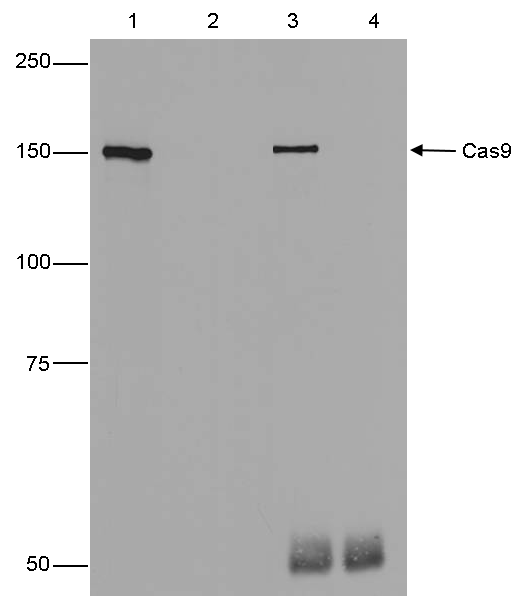
Figure 4. IP using the Diagenode antibody directed against Cas9
IP was performed on whole cell extracts (500 µg) from HeLa cells transfected with a Cas9 expression vector (lane 1 and 3), or untransfected cells (lane 2 and 4) using 1 µl of the Diagenode antibody against Cas9 (cat. No. C15310258). The immunoprecipitated proteins were subsequently analysed by Western blot. Lane 3 and 4 show the result of the IP, the input (25 µg) is shown in lane 1 and 2.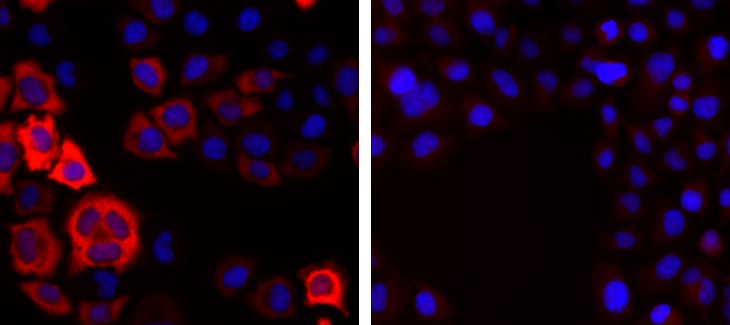
Figure 5. Immunofluorescence using the Diagenode antibody directed against Cas9
HeLa cells expressing Cas9 under the control of the tight TRE promoter were fixed in methanol at -20°C, permeabilized with acetone at -20°C and blocked with PBS containing 2% BSA. The cells were stained with the Cas9 antibody (cat. No. C15310258) diluted 1:1000, followed by incubation with a goat anti-rabbit secondary antibody coupled to AF594. Nuclei were counter-stained with Hoechst 33342. Figure 5 shows the result in the presence (left) or absence (right) of doxycycline. - お客様の声
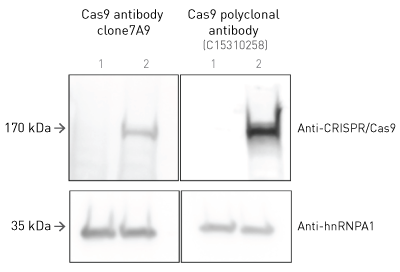
Diagenode’s CRISPR/Cas9 polyclonal antibody shows superior signal than the original clone 7A9: a researcher from EPFL in Lausanne, Switzerland has compared these two antibodies in Western blot.

Western blot was performed using HCT116 DKO cells transduced with Krab-dCas9 (2) or non-transduced (1) cells. Then, 100,000 cells were lysed in sample buffer 2x and boiled 5 min at 95°C before loading in a 15% acrylamide gel. The same sample was loaded 3x in the same gel. The membrane was cut in 3 parts for each antibody. Membrane was blocked 1h with 3% BSA at RT. Antibodies were diluted 1:1,000 in 3% BSA and incubated overnight at 4°C. Secondary incubation was done for 1h at RT (1:10,000 dilution). Anti-hnRNPA1 was used as a loading control.
EPFL in Lausanne - 出版物

How to properly cite our product/service in your work
We strongly recommend using this: CRISPR/Cas9 polyclonal antibody (Hologic Diagenode Cat# C15310258-100 Lot# A2508-004). Click here to copy to clipboard.
Using our products or services in your publication? Let us know!
Transcriptional stress induces the overexpression of novel lncRNAs that regulate the BRCA1 locus
Samantha Cruz-Ruiz et al.
Long non-coding RNAs (lncRNAs) have been shown to play a role during transcriptional regulation in response to stress. However, their function under stress caused by transcriptional inhibition has not yet been addressed. Using genome-wide assays to elucidate the transcriptional response in human cells caused by RNA ...TurboCas: A method for locus-specific labeling of genomic regions and isolating their associated protein interactome
Bercin K Cenik et al.
Regulation of gene expression during development and stress response requires the concerted action of transcription factors and chromatin-binding proteins. Because this process is cell-type specific and varies with cellular conditions, mapping of chromatin factors at individual regulatory loci is crucial for underst...Human induced pluripotent stem cells for live cell cycle monitoring and endogenous gene activation
Kim R. et al.
The fluorescence ubiquitination cell cycle inhibitor (FUCCI) has been introduced to monitor cell cycle activity in living cells, including human induced pluripotent stem cells (hiPSC) and derived cell types. We have recently developed hiPSC with stable expression of dCas9VPR for endogenous gene activation and a Citr...Methyltransferase Inhibition Enables Tgf Driven Induction of and in Cancer Cells.
Liu Y-T et al.
deletion or silencing is common across human cancer, reinforcing the general importance of bypassing its tumor suppression in cancer formation or progression. In rhabdomyosarcoma (RMS) and neuroblastoma, two common childhood cancers, the three transcripts are independently expressed to varying degrees, but one, is u...Massively parallel multi-target CRISPR system interrogates Cas9-basedtarget recognition, DNA cleavage, and DNA repair
Zou Roger S. et al.
CRISPR-Cas9 nucleases, and particularly Streptococcus pyogenes Cas9, are widespread tools for genome editing. However, many aspects of intracellular Cas9 activity and the ensuing DNA damage response remain incompletely characterized. In order to address these issues, we developed a multiplexed CRISPR approach, where...Analysis of estrogen-regulated enhancer RNAs identifies a functionalmotif required for enhancer assembly and gene expression.
Hou Tim Y and Kraus W Lee
To better understand the functions of non-coding enhancer RNAs (eRNAs), we annotated the estrogen-regulated eRNA transcriptome in estrogen receptor α (ERα)-positive breast cancer cells using PRO-cap and RNA sequencing. We then cloned a subset of the eRNAs identified, fused them to single guide RNAs, and ...Avian influenza viruses suppress innate immunity by inducingtrans-transcriptional readthrough via SSU72.
Zhao Y. et al.
Innate immunity plays critical antiviral roles. The highly virulent avian influenza viruses (AIVs) H5N1, H7N9, and H5N6 can better escape host innate immune responses than the less virulent seasonal H1N1 virus. Here, we report a mechanism by which transcriptional readthrough (TRT)-mediated suppression of innate immu...Antisense non-coding transcription represses the <i>PHO5</i> model gene<i>via</i> remodelling of promoter chromatin structure
Novačić A. et al.
Pervasive transcription of eukaryotic genomes generates non-coding transcripts with regulatory potential. We examined the effects of non-coding antisense transcription on the regulation of expression of the yeast PHO5 gene, a paradigmatic case for gene regulation through promoter chromatin remodeling. By enhancing o...A predominant enhancer co-amplified with the oncogene is necessary andsufficient for its expression in squamous cancer
Liu Y. et al.
Amplification and overexpression of the SOX2 oncogene represent a hallmark of squamous cancers originating from diverse tissue types. Here, we find that squamous cancers selectively amplify a 3’ noncoding region together with SOX2, which harbors squamous cancer-specific chromatin accessible regions. We identif...Establishment of a second generation homozygous CRISPRa human inducedpluripotent stem cell (hiPSC) line for enhanced levels of endogenous geneactivation.
Schoger Eric et al.
CRISPR/Cas9 technology based on nuclease inactive dCas9 and fused to the heterotrimeric VPR transcriptional activator is a powerful tool to enhance endogenous transcription by targeting defined genomic loci. We generated homozygous human induced pluripotent stem cell (hiPSC) lines carrying dCas9 fused to VPR along w...Establishment of two homozygous CRISPR interference (CRISPRi)knock-in human induced pluripotent stem cell (hiPSC) lines for titratableendogenous gene repression.
Schoger Eric et al.
Using nuclease-deficient dead (d)Cas9 without enzymatic activity fused to transcriptional inhibitors (CRISPRi) allows for transcriptional interference and results in a powerful tool for the elucidation of developmental, homeostatic and disease mechanisms. We inserted dCas9KRAB (CRISPRi) cassette into the AAVS1 locus...TGFβ promotes widespread enhancer chromatin opening and operates ongenomic regulatory domains.
Guerrero-Martínez J. et al.
The Transforming Growth Factor-β (TGFβ) signaling pathway controls transcription by regulating enhancer activity. How TGFβ-regulated enhancers are selected and what chromatin changes are associated with TGFβ-dependent enhancers regulation are still unclear. Here we report that TGFβ treatment...Development of an ObLiGaRe Doxycycline Inducible Cas9 system for pre-clinical cancer drug discovery.
Lundin, Anders and Porritt, Michelle J and Jaiswal, Himjyot and Seeliger, Frank and Johansson, Camilla and Bidar, Abdel Wahad and Badertscher, Lukas and Wimberger, Sandra and Davies, Emma J and Hardaker, Elizabeth and Martins, Carla P and James, Emily and
The CRISPR-Cas9 system has increased the speed and precision of genetic editing in cells and animals. However, model generation for drug development is still expensive and time-consuming, demanding more target flexibility and faster turnaround times with high reproducibility. The generation of a tightly controlled O...A gene therapy for inherited blindness using dCas9-VPR–mediatedtranscriptional activation
Böhm, Sybille and Splith, Victoria and Riedmayr, Lisa Maria and Rötzer,René Dominik and Gasparoni, Gilles and Nordström, Karl J. V. and Wagner,Johanna Elisabeth and Hinrichsmeyer, Klara Sonnie and Walter, Jörn andWahl-Schott, Christian and Fenske, Stef
Catalytically inactive dCas9 fused to transcriptional activators (dCas9-VPR) enables activation of silent genes. Many disease genes have counterparts, which serve similar functions but are expressed in distinct cell types. One attractive option to compensate for the missing function of a defective gene could be to t...Guidelines for optimized gene knockout using CRISPR/Cas9
Campenhout CV et al.
CRISPR/Cas9 technology has evolved as the most powerful approach to generate genetic models both for fundamental and preclinical research. Despite its apparent simplicity, the outcome of a genome-editing experiment can be substantially impacted by technical parameters and biological considerations. Here, we present ...CRISPR-mediated activation of a promoter or enhancer rescues obesity caused by haploinsufficiency.
Matharu N, Rattanasopha S, Tamura S, Maliskova L, Wang Y, Bernard A, Hardin A, Eckalbar WL, Vaisse C, Ahituv N
A wide range of human diseases result from haploinsufficiency, where the function of one of the two gene copies is lost. Here, we targeted the remaining functional copy of a haploinsufficient gene using CRISPR-mediated activation (CRISPRa) in and heterozygous mouse models to rescue their obesity phenotype. Transgeni...(Po)STAC (Polycistronic SunTAg modified CRISPR) enables live-cell and fixed-cell super-resolution imaging of multiple genes
Neguembor M.V. et al.
CRISPR/dCas9-based labeling has allowed direct visualization of genomic regions in living cells. However, poor labeling efficiency and signal-to-background ratio have limited its application to visualize genome organization using super-resolution microscopy. We developed (Po)STAC (Polycistronic SunTAg modified CRISP...A self-inactivating system for AAV-mediated in vivo base editing
Zuo Y. et al.
DNA base editors have been harnessed as an exciting therapeutic platform for human diseases and are rapidly progressing into human clinical trials. However, persistent expression of base editors delivered via adeno-associated virus (AAV) poses concerns with specificity and immunogenicity. Here we develop selfinactiv...